Capítulo XVII: El misterio de la vida.
En el capítulo anterior nos encontrábamos a las puertas de enfrentarnos cara a cara con el misterio del origen de la vida. Para los lectores que se incorporen ahora o los que tengan la memoria frágil, recordemos las paradojas que se nos habían presentado hasta ahora a la hora de encarar este enigma y como se habían resuelto:
Paradoja del Huevo y la Gallina: Preguntábamos qué fue primero, si los mecanismos de reproducción del ADN o la información contenida en el ADN capaz de generar y coordinar dichos mecanismos (de caracter enzimático). La solución la encontramos en el ARN, una molécula algo más sencilla capaz en principio de replicarse por si misma, sin ayuda de nada ni nadie. El ARN habría ido desarrollándose hasta actuar como los cimientos para la aparición del ADN (recordemos que en la actualidad sigue ocupando un muy importante papel auxiliar).
Paradoja de la aparición de secuencias funcionales: El ARN, dispuesto a guisarse y comerse el solo su propia replicación, estaría expuesto a la posibilidad de sufrir numerosos errores al ensamblar dichas réplicas, dando al traste con todo en muy pocas replicaciones. Sin embargo se descubrió que ciertas combinaciones de secuencias largas de nucleótidos (los eslabones que componen el ADN y el ARN) podían favorecer la efectividad de aquellas que fueran correctas frente a las que tuvieran errores, salvando de este modo la perpetuación de la información necesaria para seguir con la replicación (y por lo tanto con la evolución de la vida).
Sin embargo, he aquí que superadas estas dificultades aparecieron otras dos, también en forma de paradoja. Repasémoslas y tratemos de resolverlas.
La Paradoja del agua: Todos hemos escuchado una y mil veces lo importante que es el agua para la vida, y no es ninguna tontería. El agua es el medio perfecto para que, disueltas en ella, se puedan acumular distintas sustancias de modo que puedan reaccionar unas con otras, lo cual en otras circunstancias sería muy difícil. Si no estuviera ahí el líquido elemento para hacer de "tablero de juego" para la química de la vida, no estaríamos aquí. Diversos investigadores han tratado de pensar en otras sustancias que pudieran cumplir un papel similar al del agua, pero ninguna se mantiene líquida en una gama tan amplia de temperaturas y presiones, y lo más importante, en ninguna otra el estado sólido es menos denso que el líquido. Todos estamos acostumbrados a ver flotar el hielo en agua, pero si no fuera así y se hundiera no aislaría el calor del agua que tuviera debajo (como hace por ejemplo en el ártico), con lo cual dicho calor se perdería generando un bucle de enfriamiento que terminaría por convertir todo en hielo. Gracias a ello nuestros océanos no se encuentran actualmente congelados. Pero situados en la Tierra primitiva, todo esto no era tan bonito. El agua es un gran disolvente, y de manera natural tiende a disgregar las cadenas de nucleótidos que veíamos conforman el ADN y el ARN. Así pues... ¿como pudieron estas cadenas formarse, sobre todo las del ARN ancestral, frente a un handicap acuoso tan severo? De algún modo necesitamos un mecanismo que favorezca el ensamblaje de nucleótidos y otras moléculas orgánicas compensando su tendencia separarse por culpa del agua.
Graham Cairns-Smith, de la Universidad de Glasgow, elaboró una teoría muy arriesgada para explicar el improbable nacimiento de la vida bioquímica, especulando con que esta en realidad fuera heredera de una vida mineral precedente. Puedo imaginar el rostro estupefacto del lector al verse enfrentado a la posibilidad de formas de vida minerales, pero una vez explicado en seguida verá como esto no es tan raro como parece. Resulta que ciertos cristales de arcilla al formarse no son ni perfectos ni uniformes sino que contienen defectos, y dichos defectos se extienden según lo hace el cristal, y cuando este se fractura los distintos fragmentos conservan el patrón defectuoso original. De este modo podemos tener distintos tipos de cristales cada uno con distintas configuraciones, que se reproducirían necesariamente a distintas velocidades, de manera que aquellos cristales "más eficaces" tenderían a hacerse mayoritarios, solo para verse sustituidos por el siguiente patrón que apareciera con aún mayor "eficacia reproductiva". En la practica no estaríamos ante otra cosa que una selección natural de cristales de arcilla, que se dirigiría hacia una cada vez mayor complejidad solo para poder seguir haciendo más y mejores copias de si mismos, exactamente como lo hace la vida.
Maravilloso y a la vez perturbador este panorama, ¿pero en que momento se podría haber dado el salto de la vida mineral a la vida bioquímica? Aquí es donde esta teoría pisa un terreno menos firme. Graham Cairns-Smith apuntaba que los cristales de arcilla podrían incluir en sus estructuras a ciertas moléculas orgánicas (precursoras del ARN) que les ayudarían en la gestión de su crecimiento y cada vez mayor complejidad. Finalmente la vida bioquímica "tomaría el control" de la situación, dejando atrás a los pobres cristales de arcilla y alcanzando nuevas cotas de éxito. Esta teoría nos ayudaría a eludir la laguna previa a la aparición del ARN, pero sigue sin estar claro como se produciría el relevo del "poder genético" de los cristales de arcilla a las moléculas orgánicas, y ni el señor Cairns-Smith ni nadie ha realizado experimentos que ayuden a arrojar luz sobre esto.
Sin embargo, cada vez se sospecha más que después de todo ciertas superficies minerales (de arcilla o de otros materiales) desempeñaron un papel crucial en el nacimiento de la vida. Estas superficies, al contar con capacidad de absorción de pequeñas moléculas orgánicas, pudieron funcionar como unos potentes catalizadores, ayudando a que dichas moléculas se juntaran dentro de espacios reducidos (como en una especie de local de citas) y tuvieran allí lugar reacciones químicas que en otro lugar hubiera sido muy difícil que se produjeran. Para imaginarlo tan solo comparen un escenario pequeño y casi bidimensional sobre una superficie arcillosa con el enorme y hostil mundo tridimensional de una disolución acuosa.
Otra teoría muy prometedora es la que desarrolló en su día el científico soviético Alexander Oparin. Tratando de pensar en como sustancias abióticas (no vivas) podían combinarse generando vida, dio con la teoría de los protobiontes, es decir, agregados de moléculas no vivos que habrían podido evolucionar hasta dar lugar a la primera criatura sí viva. Pronto se vio que no era tan difícil imaginar e incluso generar en laboratorio dichos eslabones perdidos entre lo inerte y lo orgánico. Hablamos de los protobiontes. Básicamente son burbujas de grasa que cuentan con cierta facilidad para formarse dadas las condiciones adecuadas, las cuales dicho sea de paso se daban en la Tierra primitiva. Dicho así parece algo muy tosco y alejado de nada remotamente parecido a un ser vivo, pero en realidad la estructura exterior de nuestras células es en principio la misma que la de los protobiontes. Todas las membranas celulares y también las de algunos virus están formada por una bicapalipídica, la cual como su nombre índica está constituida por dos capas de fosfolípidos (un tipo de molécula de grasa) cada una de las cuales cuenta con dos extremos con propiedades opuestas. Un extremo es hidrófobo, es decir, que es repelido por el agua, y el otro hidrófilo, con justo el comportamiento contrario. Así pues, cuando arrojamos un puñado de fosfolípidos al agua todos estarán tratando de separarse del agua por un lado y de juntarse con ella por el otro, lo cual en la práctica lleva a que se junten en una espera de doble capa, con los extremos hidrófilos apuntando hacia fuera y hacia dentro de la misma, y los extremos hidrófobos unidos y protegidos del agua entre ambas capas. La imagen que dejo a continuación ayudará al lector a visualizarlo.
 |
| Bicapalipídica. |
Pues bien, es en teoría posible que, aisladas dentro de estos protobiontes con forma de burbujas de grasa quedasen encerrados cierto número de moléculas orgánicas, que en estas “circunstancias de intimidad” lo tendrían fácil para interactuar unas con otras como en el caso de la arcilla, y más aún si entre las mismas hubiera enzimas que ayudaran a hacer "la fiesta más divertida". A este tipo de protobiontes Oparin los llamó coacervados y él mismo y los investigadores que le siguieron han logrado generarlos en laboratorio y estudiarlos. En concreto algunos coacervados formados dentro de soluciones ricas en proteínas tienen la capacidad de crecer incorporando diversas moléculas orgánicas a su composición hasta que llegado un momento alcanzan un desequilibrio químico en su interior, tras lo cual se rompen generando "gotitas" que darán lugar a nuevos coacervados destinados a repetir la misma secuencia de acciones. Tal vez de algún modo la selección natural pudiera haberse colado en este proceso, por ejemplo si los coacervados resultantes de la destrucción de otro pudieran heredar de algún modo la composición química de su predecesor. El wikipedista anómimo que ha tratado sobre estos temas en el correspondiente artículo de la Wikipedia, nos lanza esta teoría:
Se forma una molécula organizada y relativamente estable.
1) Con el tiempo otros materiales que corresponden a su conformación a una segunda molécula complementaria, es decir una macromolécula que se forma en el coacervado enlazando al mismo y creando algo que se puede llamar como un molde químico.
2) Este molde se separa del coacervado.
3)Debido a la gran cantidad de compuestos orgánicos que hay en el medio, el molde comienza a atraer compuestos que puede enlazar, recreando el coacervado original.
4) El molde se separa (con su probable destrucción) del coacervado así dando lugar a un "hijo".
Podría ser que los coacervados evolucionaran hasta generar formas vivas de esta manera, aunque dentro de unos pocos párrafos descubriremos que esto no es tan fácil después de todo.
Hay muchas más teorías que nos hablan de como pudieron juntarse y reaccionar diversas moléculas orgánicas, en especial nucleótidos para formar cadenas, paradojas del agua aparte. Otra de ellas se plantea ciertos desequilibrios termodinámicos como la "inestabilidad de Bernard", que podrían dar lugar a comportamientos bioquímicos muy complejos, pero para poderles explicar esto primero debería de entenderlo y no es el caso, no obstante me remito a la bibliografía para el lector que quiera saber más.
Pero incluso asumiendo que en la Tierra primitiva pudiera disponerse de los medios adecuados para para ello (sea mediante coacervados, seres de arcilla o carambolas termodinámicas), no estamos hablando de generar una cadena de nucleótidos cualquiera, tratamos en realidad de ver como apareció algo parecido al ARN, una cadena de nucleótidos dotada de un tamaño y una complejidad muy considerables, tanto como para poder replicarse a si misma de modo autónomo. Este es desde luego el quid de la cuestión y con ello nos damos de bruces con nuestra paradoja final:
Como decíamos, en varias ocasiones algunos de los más brillantes científicos se han propuesto la tarea de juntar todos los ingredientes y circunstancias conocidos favorables a la síntesis de vida solo para ver que pasa. Y por más empeño que han puesto en ello y más "empujoncitos" que han podido darle a vida para que aparezca dentro de un tubo de ensayo, hasta ahora solo se ha conseguido generar una especie de limo con una apariencia similar a la del alquitrán. En 1953, siguiendo las ideas que se tenían en la época sobre las condiciones de la Tierra primitiva y tratando de ir un poco más lejos que Alexander Oparin, el científico americano Stanley Miller protagonizó un experimento que pasaría a los anales de la historia. Simuló la atmósfera primordial de nuestro planeta, le suministró energía en forma de pequeños relámpagos (muy al estilo Victor Frankenstein) y... ¿que obtuvo? Pues logró sacar varios aminoácidos entre otras varias moléculas orgánicas complejas. También se formaron protobiontes.
 |
| Stanley Miller y su experimento. |
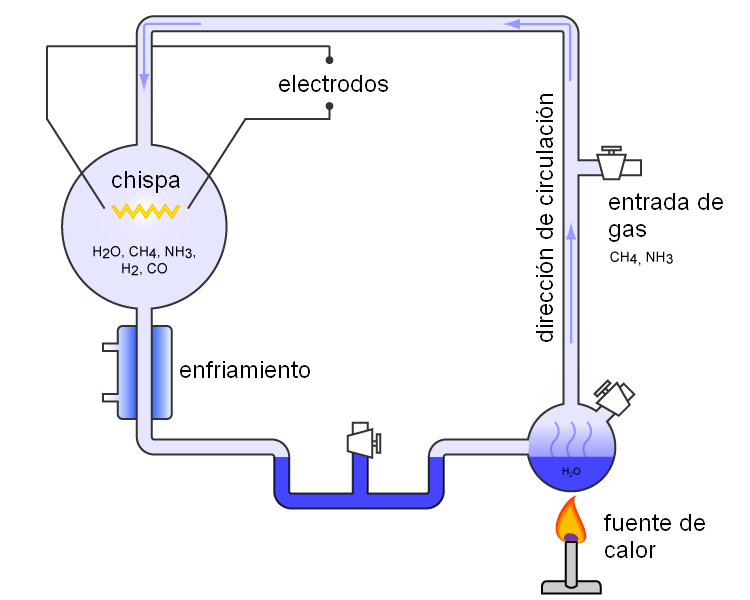 |
| Esquema del experimento de Stanley Miller. |
Sin embargo, las condiciones que se planteó Miller eran mucho más optimistas que las que hoy pensamos que tenían lugar realmente en los primeros tiempos de nuestro planeta. Ahora sabemos que había trazas de oxigeno, un gas venenoso para la vida primitiva, y que la presencia de un "rico caldo de cultivo" de sustancias orgánicas prestas a combinarse de mil maneras no era tan abundante ni tan variado. Además, nuestro limo alquitranoso de aminoácidos y nucleótidos sueltos (obtenido en laboratorio en condiciones muy benignas) está tan lejos de una molécula de ARN y no digamos ya de ADN como yo de viajar a Marte. Ni siquiera se han podido sintetizar proteínas juntando todos los ingredientes adecuados para ello. Entonces... ¿qué?
El astrofísico Fred Hoyle, escéptico sobre el origen azaroso de la vida, nos hace cálculos como el siguiente: "la probabilidad de que una cadena proteica de 100 aminoácidos – formada a partir de los 20 tipos con los que se construyen las proteínas - tenga exactamente la secuencia de una enzima (1) actual de esa longitud es de uno entre 20 elevado a 100". Cojan su calculadora científica y echen números. Resulta que aunque toda la faz de la Tierra hubiera estado cubierta desde sus orígenes por una capa de un metro de grosor de aminoácidos combinándose al azar cada milésima de segundo, sería tremendamente improbable que hubiera surgido por azar nuestra enzima. Y la cosa se pondría todavía peor para la aparición del ARN autoreplicante a partir de combinaciones aleatorias de nucleótidos. Fred Hoyle argumenta a favor de una Inteligencia universal (bonito eufemismo para hablar de una deidad) que habría amañado las cuentas a favor de la vida. No sé ustedes, pero a mi me parece una solución muy burda, del estilo de "ah, eso... pues... ¡vino un mago y lo hizo!". Si les parece mejor intentemos hilar más fino en pos de la solución más consistente para este enigma.
Resulta que, como veíamos el otro día con "el juego de la vida", ciertos sistemas tienden espontáneamente a la autorganización. Ejemplos de ello nos rodean por doquier. Por ejemplo, en el Universo recién nacido a partir de una convulsa sopa de partículas elementales y radiación surgieron complejos y elaborados átomos, cada uno con sus propiedades específicas, y después a partir de ellos casi infinitas combinaciones de moléculas que danzan unas con otras en elegantes coreografías químicas. También se han descubierto maravillosos patrones matemáticos que simplemente surgen espontáneamente del azar, como por ejemplo el Conjunto Mandelbrot, cuya imagen y explicación dejo a continuación.
 |
| Esta bella imagen, ampliada suficientes veces se contiene a si misma en cada uno de sus pequeños detalles (que surgen a modo de pelillos), los cuales a su vez se contienen en ellos a sí mismos y así infinitamente, como una especie de muñecas rusas sin fin. Se trata de la proyección de un tipo concreto de ecuación compleja sobre un plano (llamado en consecuencia plano complejo). Para saber más: http://es.wikipedia.org/wiki/Conjunto_de_Mandelbrot |
Según Fred Hoyle, el puro y duro azar nunca debería de habernos dejado ir más allá de la sopa inicial de partículas elementales y radiación que generó el Big Bang, ni tampoco permitiría las elegantes formas matemáticas que hemos visto. Antonio Lima de Faria, de la Univeridad de Lund, nos habla de "autoevolución", un tipo de evolución que no necesitaría de inteligencia o de genes para ser prodigiosamente efectiva. Según él, "la evolución es un fenómeno inherente a la materia y a la energía" y "cada nivel es prisionero del precedente, estando la propia evolución biológica totalmente condicionada por el orden de las evoluciones previas, la de las partículas elementales, la de los elementos químicos, la de los minerales, etc.". Incluso experimentos químicos clásicos demuestran que los aminoácidos, dejados a su "libre albedrío" no se combinan de cualquier manera, sino que se forman espontáneamente unas secuencias con alta probabilidad mientras que otras no aparecen. Incluso esos aminoácidos engarzados al azar muestran habilidades enzimáticas débiles de diverso tipo.
Vale vale, pero podemos imaginarnos a Hoyle dispuesto a frenar nuestro optimismo aludiendo a la selección natural como único mecanismo espontáneo creador de orden en lo que a la vida se refiere. Y la selección natural es una diosa ciega y caprichosa que actúa al azar. Por fortuna para nuestra búsqueda de una explicación científicamente explicable del origen de la vida, tenemos un último as guardado bajo la manga. Se trata de los últimos avances en genética molecular, concretamente en un campo de estudio que ha venido a llamarse "combinatoria genética".
Resulta
que los genes de los eucariotas, los organismos compuestos del tipo
de célula más avanzada (la célula eucariota, véase nota 2), están
compuestos de "trozos". Por un lado los "exones",
segmentos que codifican proteínas, y luego los "intrones",
otros fragmentos que no codifican nada y que están ahí
aparentemente de relleno (si uno ve una película en la TV, los
intrones serían como los anuncios). Por un momento se denominó al
ADN intrónico como "ADN basura", porque se pensó que se
podía prescindir de él y en realidad no era más que un desecho de
recombinaciones pasadas que no había podido ser eliminado. Sin
embargo más del 90% de nuestro ADN esta formado por intrones, lo
cual constituye uno de los más grandes misterios de la biología
moderna. Recientes investigaciones han detectado que si bien los
intrones son eliminados por el ARN a la hora de copiar y manipular la
información genética para sintetizar proteínas, estos parecen
desempeñar funciones más sutiles, por ejemplo influyendo sobre
algunas proteínas que tienen la capacidad de unirse al ADN con el
fin de realizar ciertas funciones de transcripción y replicación.
Pero a efectos de tratar de discernir el origen de la complejidad de
la vida, descubrimos una ventaja sorprendente de los intrones:
dispersan la información útil para la expresión de las proteínas,
lo cual protege a la misma de posibles mutaciones surgidas al azar
(como ya vimos) y además permite recombinaciones novedosas de
exones, sacando a la luz montajes inéditos. Dicho de otro modo, es
más fácil conseguir una proteína nueva mediante combinaciones de
exones gracias a la variabilidad añadida por los intrones que
mediante a mutaciones sucesivas de una proteína inicial. Ello habría
acelerado enormemente la evolución de la vida. Apoya esta teoría el
que hoy en día sabemos que muchos exones se corresponden con
unidades estructurales discretas de las proteínas, llamados
"dominios". Es más. Se conocen casos en los que un mismo
dominio, con ligeros retoques, puede construir proteínas de diversa
función. Para ayudar al lector a verlo, la evolución puede jugar
de este modo con una construcción modular de multitud de proteínas
partiendo de un número muy limitado de módulos "en catálogo".
Algo así como montar un mueble de Ikea sin tener que fabricar
previamente las piezas.
Muchas
veces ha tratado de argumentarse en contra de la utilidad de los
intrones el hecho de que las bacterias, organismos de gran éxito
evolutivo, carecen de ellos. Sin embargo las bacterias en algún
momento de su historia debieron de sacrificar su complejidad y la
posibilidad de potenciar su evolución en pos de una mayor eficacia a
la hora de reproducirse: cuantas más cortas son las secuencias, más
rápidamente y con menos gasto se copian. Gracias a ello las
bacterias pueden proliferar del modo vertiginoso que lo hacen.
Además, las bacterias han desarrollado métodos alternativos para
tratar de ganar variabilidad genética, como el libre intercambio de
genes entre ellas, algo que ya veremos en el próximo capítulo.
Estos
mecanismos de "diversidad compleja" que hemos estado
bosquejeando explicarían por ejemplo porqué la vida surgió tan
rápido mientras que luego la selección natural, con todo su
supuesto poder, tardó unos 2.000 millones de años en dar con las células eucariotas, es decir, con las células modernas con núcleo que
componen a animales, plantas y hongos.
Algún
escéptico podría alegar que, pese a todo esto, sigue siendo muy
difícil el desarrollo de los sistemas de traducción genética
modernos, recuérdese, el modo en que la información contenida en el
ADN es descifrada y utilizada para fabricar las proteínas con las
que funcionamos.
Una
posible explicación es la llamada teoría del mecanismo secuencial.
Un ejemplo: queremos abrir una caja fuerte con una combinación de 6
cifras. Probando al azar y tardando un segundo por cada prueba,
necesitaríamos más de cinco días de esfuerzos consecutivos para
abrirla. Pero si la caja tuviera seis cerrojos consecutivos con una
combinación de una cifra cada uno, solo necesitaríamos un minuto.
Tal vez la traducción genética fue "abriendo cerrojos".
Imaginen que una arcaica forma de vida consigue una pequeña ventaja
evolutiva con un sistema de traducción incompleto. Dicha ventaja se
seleccionaría y guardaría hasta que se presentase otra y así
sucesivamente. Todo esto aún no ha podido ser probado, pero ofrece
una salida a como mecanismos tan complejos podrían haberse
desarrollado de modo azaroso en relativamente pocos millones de años.
Hay
no obstante otro acercamiento a este problema con el que terminaremos
este capítulo y que nos acercará al origen mismo de la vida, o al
menos del código genético en el que la vida moderna se basa. No
perdamos de vista que nosotros y toda nuestra civilización podríamos
en realidad ser considerados tan solo como una especie de sistema externo
de memoria usado por nuestros genes para sus propios intereses (3).
Pero veamos al fin como pudo empezar todo esto.
Recordemos
que los nucleótidos del ADN (A, U, G y C) se organizan en tripletes
para, en función de distintas combinaciones, codificar proteínas.
ACU, GAC, CGU, etc funcionan como palabras con las cuales se escriben
las instrucciones necesarias para el ensamblaje de los 20 tipos
distintos de aminoácidos que componen las mencionadas proteínas.
Bien, pero... ¿que hay detrás de este código de ensamblaje? ¿y
porque solo se usan 20 tipos de aminoácidos cuando sabemos que existen
más? Puesto que estos dos hechos quedaron establecidos en el momento
en que la vida, apareciera como apareciera, quedó definida como lo
es hoy en día, podemos examinar la huella que pudiera quedar en
ellos acerca de qué fue lo que pasó.
Empecemos
con el lenguaje genético. Los tripletes que antes veíamos aparecen
"escritos" todos
juntos, sin puntos de separación ni nada parecido (CAAGUAUGCG...).
Actualmente las células (y nuestros expertos en ingeniería
genética) saben desde donde empezar a leer, y a partir de ahí solo
hay que ir contando de tres en tres para desencriptar el mensaje
(CAA-GUA-GCG-...). Sin embargo se sabe que las células primitivas
contaban con una genética más rudimentaria que no hacía fácil
saber donde empezar a leer el mensaje para encontrarle sentido, con
lo cual los biólogos se han partido la cabeza tratando de buscar
sistemas de puntuación ancestrales. Por ejemplo, un cierto triplete
podría haber marcado el comienzo de la información útil y otro el
final, de modo parecido a nuestros espacios entre palabras. Solo
algún tiempo después, la mejora en los sistemas de traducción del
mensaje genético de las células modernas habría hecho que no fuera
necesaria puntuación y se habría ganado más eficacia y riqueza de
vocabulario. Sin embargo de ser esto así debería de quedar todavía
un rastro de los arcanos símbolos de puntuación. Y esto es lo que
en efecto sucede. Los tripletes más cargados de significado siguen
ocupando la posición central entre otros más accesorios o incluso
vacíos en apariencia de sentido.
Este
hecho nos lleva a una conclusión enigmática: desde el principio la
vida se complicó la vida (valga la redundancia) para codificar sus
mensajes genéticos en tripletes. Sabemos que no fue primero en
cuartetos ni quintetos de nucleótidos, porque no se podrían haber
reducido después a paquetes de tres. Piensen a modo de ejemplo en
HACE RATO COMI BIEN PERO POCO, una información que según el
contexto puede ser muy útil. Si tratamos de reducirlo a tripletes de
letras, nos encontraríamos con HAC EGR ANR ATO COM IBI ENP ERO POC
O, expresión que como poco nos valdría una mirada de desconfianza y
extrañeza por parte de nuestro contertulio. Fueron tripletes
entonces desde el principio. ¿Pero porqué? Este era un misterio
hasta que hace relativamente poco tiempo estudios en laboratorio empezaron
a encontrar ciertas afinidades químicas entre tripletes de
nucleótidos y aminoácidos. Parece ser que la química de la vida
primitiva podría haber amañado un poco las cuentas usando los
tripletes adecuados, lo cual habría ahorrado un ingente esfuerzo de
azarosas pruebas y errores, facilitando sobre manera el origen de los
mecanismos de expresión genética actuales, que líneas más arriba
tan azarosamente improbables nos parecían. Y más aún, algunos
aminoácidos serían más afines que otros, con lo que solo un
selecto grupo de ellos se irían seleccionando, hasta quedarse en los
20 actuales, que como hemos dicho son los únicos utilizados por toda
la vida conocida. ¿Por qué no más de 20? Porque parece ser que
alcanzada esa cifra los costes de sintetizar un nuevo tipo de
aminoácido excederían a los beneficios.
De
ser esto así, los 20 aminoácidos actuales y el sistema de tripletes
de nucleótidos que los codifica serían simplemente el resultado de
un "accidente congelado" a partir de un desarrollo azaroso,
condicionado y beneficiado eso sí por las afinidades
triplete-aminoácido antes mencionadas. Según esto, nuestro código
y sistema genético no es el mejor posible, si no solamente aquel que
se desarrolló y evolucionó con más facilidad dadas unas
determinadas condiciones iniciales aleatorias. Cualquier laboratorio
de genética podría, con los recursos suficientes, diseñar un
código y un sistema mejores. Si algún día hablan con un genetista
probablemente les mostrará admiración por como funciona nuestro
ADN, pero también los errores que tiene y como se lo podría
perfeccionar (algo que ya se ha empezado a hacer por parte de algunos
ingenieros genéticos como Craig Venter).
De
nuevo buscamos nuestro origen y lo encontramos en el azar. Pero en
este caso, no es un azar cualquiera, sino que de él puede surgir de
modo natural la belleza de la vida como una propiedad emergente más
de nuestro sorprendente universo.
Como
ven, solo hemos arrojado una tenue luz sobre el hondo misterio del
origen de la vida, acaso nunca logremos resolverlo del todo, quien
sabe. No obstante no con ello terminan los enigmas que rodean a esta
extraña aventura que es la vida, más bien con él comienzan. Pues
una cosa es empezar con una célula arcaica y primitiva y otra muy
distinta terminar con criaturas como elefantes, geranios, champiñones
y seres humanos. ¿Que creemos saber sobre la evolución de la vida?
En las últimas décadas se han tratado de estirar y forzar los
mecanismos de la selección natural para explicarlo, pero
poco a poco resulta más evidente que esta teoría, sin ser
incorrecta, no es suficiente. Se necesita alguna otra que sea más
amplia y poderosa para poder explicar los desafíos que nos plantea
la evolución de la vida, y de ello nos ocuparemos, más ello será
en el siguiente capítulo.
Bibliografía:
- Texto en el que me he basado / he copiado principalmente: "Luces y sombras sobre el origen de la vida". Juan Antonio Aguilera Mochón, departamento de Bioquímica Molecular de la Universidad de Granada.
- "La Nueva Mente del Emperador", Roger Penrose, editorial Debolsillo.
Notas:
1) Recordemos
que las enzimas son un tipo de proteína que usa el ADN para poder
facilitar y regular sus funciones.
2) Las
células modernas, que al contrario que bacterias y arqueo bacterias
(los otros dos dominios de la vida) poseen un núcleo que almacena
la información genética así como algunos otros orgánulos que les
garantizan una mayor complejidad y eficiencia. Las células
eucariotas componen a animales, plantas y hongos, así como a los
protistas (organismos unicelulares como las amebas y los
paramecios).
3) Léase
el libro de Richard Dawkins “El gen egoísta”.
No hay comentarios:
Publicar un comentario